LABORATORY OF NUTRITIONAL BIOCHEMISTRY,
SCHOOL OF FOOD AND NUTRITIONAL SCIENCES,
UNIVERSITY OF SHIZUOKA
SCHOOL OF FOOD AND NUTRITIONAL SCIENCES,
UNIVERSITY OF SHIZUOKA
運動トレーニングによる骨格筋での代謝変化の分子メカニズム
転写共役因子peroxisome proliferators-activated receptor-γ co-activator-1α(PGC-1α)は、核内受容体や転写因子と複合体を形成し遺伝子の転写、特にミトコンドリア生合成に関わる遺伝子を系統的に発現制御する。持久的な運動トレーニングは、骨格筋のミトコンドリア量増加、筋線維タイプ変化(赤筋化)、毛細血管新生などをもたらし、エネルギー代謝を促進する。我々の研究を含めた最近の知見では、運動トレーニングにより骨格筋にもたらされる多様な変化の多くが、PGC-1αの発現増加が引き金になって生じていることが判明している。
我々はこれまでに、運動による骨格筋PGC-1α発現増加にはβ2アドレナリン受容体刺激が必要なこと(Endocrinology 2007)、PGC-1αに3種のアイソフォームが存在し、各アイソフォームの発現やその調節機序は臓器ごとに異なることなどを明らかにしてきた(Endocrinology 2008, Am J Physiol Endocrinol Metab 2011)(図1)。また各PGC-1αアイソフォームを骨格筋特異的に過剰発現させたトランスジェニックマウスを独自に作製した(J Biol Chem 2003, Am J Pathol 2006, Endocrinology 2008)。さらに運動刺激に対する反応が最も高いアイソフォームであるPGC-1α-bを骨格筋特異的に過剰発現させたマウスにおいて、運動時の脂肪燃焼効率の増加、最大酸素摂取量増加、酸素利用能向上といった、持久的トレーニング効果を模する表現型を示すことを見出した(PLoS ONE 2011)(J Appl Physiol 2014)(図2)。
また、PGC-1αの増加は骨格筋でのbranched-chain aminotransferase (BCAT) 2, branched-chain α-keto acid dehydrogenase (BCKDH)の発現増加を引き起こし、分岐鎖アミノ酸代謝を促進することを明らかにした(PLoS ONE 2014)(図3)。
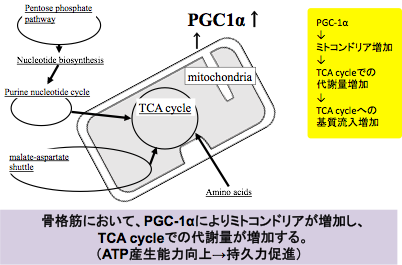
PGC1α過剰発現マウスの骨格筋を使用して網羅的に代謝産物量の変化を検出するメタボローム解析を行った。その結果、TCA回路、ペントースリン酸経路そしてBCAAやβ-アラニンを含むアミノ酸代謝が活性化していることが判明した。TCA回路、BCAAを含むアミノ酸代謝の協調的な調節はPGC1αがエネルギー代謝に重要な役割を担っていることを示している。さらに、運動時に活性化することが知られているプリンヌクレオチド回路とアスパラギン酸-リンゴ酸シャトルが活性化していた。これらの結果はPGC1αが運動時のエネルギー代謝を調節している重要な因子であることを示している(PLoS ONE 2015)(図4)。
運動後の細胞内カルシウムイオンの恒常性におけるミトコンドリアの役割を検討し,ミトコンドリアによるカルシウムイオンの取込が非常に重要であることをin vivoモデルを用いて明らかにした( Am J Physiol Regul Integr Comp Physiol 2017)。
図1
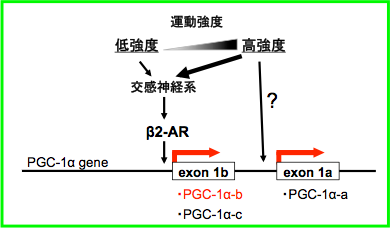

図2

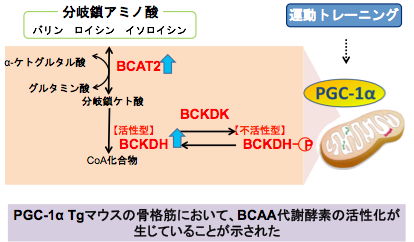
図4